Медузы в неволе
Австралийским учёным из Центра исследований рифов (CRC Reef Research Centre) впервые удалось вырастить в неволе медузу Carukia barnesi, обладающую смертоносным жалом.
Медуза-убийца на самом деле крошечная — длина её купола всего-навсего 12 миллиметров. Однако именно на это животное возлагается ответственность за возникновение синдрома Ируканджи (Irukandji syndrome), который убил в Австралии двух туристов в 2002 году.
Всё начинается с укуса, подобного комариному. В течение часа жертвы испытывают сильную боль в пояснице, прострелы по всему тела, судороги, тошноту, рвоту, обильно потеют и кашляют.
Последствия крайне серьёзны: от паралича до смерти кровоизлияния в мозг или остановки сердца.
Захваченная медуза прошла планктоническую стадию и теперь содержится в аквариуме. Добиться размножения медуз в неволе было первой стадией в развитии противоядия. В целом же потребуется изучить от 10 тысяч до миллиона медуз.
Регенерация полипов и медуз.

Процесс регенерации специфичен, т. е. из фрагмента ткани полипа образуется только полип, а из фрагмента ткани медузы - только медуза. Эта специфичность даже более узкая, те. из фрагмента ткани регенерирует полип того же типа.
Так, из участков стебля гидрактиний могут образоваться разные типы полипов: из фрагментов стебля питающих полипов - только питающие полипы, а из фрагментов репродуктивных полипов - только репродуктивные полипы.
Эти данные позволяют сделать два вывода.
1. Поскольку во взрослом функционирующем организме кишечнополостных имеются специализированные клетки (книдобласты, эпидермальные, железистые, мышечные, нервные клетки и т. д.), то при регенерации из кусочка ткани они должны возникать либо из таких же уже имеющихся клеток, либо в результате трансформации одного типа клеток в другой.
2. Фрагменты тканей, вероятно, «знают», из какого взрослого организма они произошли, и перестраиваются в соответствии с этим.
Сравните эти результаты и выводы с результатами и выводами, полученными при исследовании регенерации у ацетабулярии, описанной в седьмой главе. Обратите внимание на то, что они в основном сходны, за исключением того, что в одном случае речь идет о регенерации структуры из части, состоящей из клеток, а в другом - клеток нет, но специфическая организация существует.
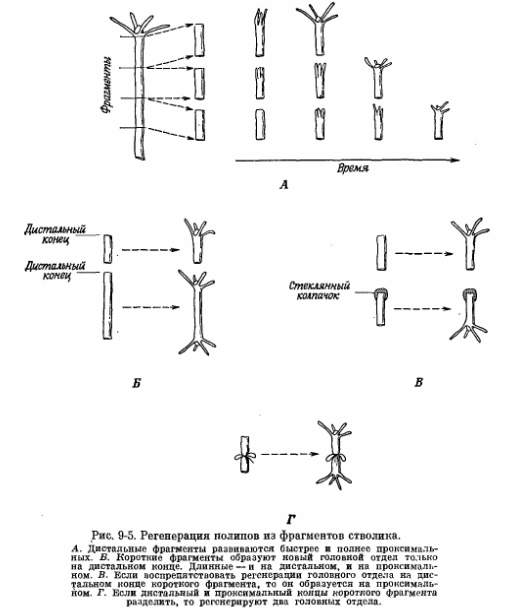
При изучении регенерации кишечнополостных из фрагментов стебля обнаружены и другие интересные особенности (рис. 9-5). Во-первых, скорость и полнота регенерации нового головного отдела зависят от того, из какой части стебля взят фрагмент. Если стебель полипа разрезать на несколько фрагментов равного размера, то фрагмент, ближе всего расположенный к головному отделу (дистальный конец), наиболее быстро и полно образует новый головной конец, а фрагмент, взятый из самой нижней части стебля (проксимальный конец),наиболее медленно. Другими словами, вдоль всего стебля существует градиент способности к образованию нового головного отдела. Кроме того, если в одном участке стебля регенерирует новый головной отдел, то его образование в другом участке подавляется. Например, на фрагменте стебля умеренной длины образуется только один новый головной отдел, причем всегда только в области разреза на дистальном конце фрагмента. Однако если воспрепятствовать регенерации головного отдела на дистальном конце, надев на него стеклянный колпачок, то головной отдел возникнет из проксимального конца (хотя медленнее, чем из дистального). Избежать действия одного конца фрагмента на другой можно двумя способами: 1) использовать достаточно длинный фрагмент ствола, чтобы оба конца находились на значительном расстоянии друг от друга, и 2) путем физиологического разобщения двух концов. Последнее можно осуществить, перевязав фрагмент ствола посередине и предотвратив тем самым перемещение веществ и клеток. При этом головные отделы регенерируют на обоих концах фрагмента, причем никаких конкурентных отношений не отмечено.

Выше на фото - перпурная медуза, Калифорния.
Это явление характерно и для других развивающихся систем. Примером может служить развитие задней конечности у куриных зародышей. Как будет описано в гл. 10, конечность возникает в виде небольшого выроста в определенном участке боковой поверхности зародыша. Показано, что окружающие участки также могут образовать конечность. Почему же конечность образуется именно там, где она образуется, и почему другие участки, из которых она может развиться, в норме не образуют конечности?
Система, в которой в определенном направлении могут развиваться многие части, но в действительности развивается только одна, называется морфогенетическим полем. Мы подробно рассмотрим эту систему в следующем разделе. Доминирование одной части поля над другой обеспечивает упорядоченность развития. Благодаря этому у полипа образуется один головной отдел именно там, где он должен быть, а у цыпленка образуются также на своих местах один хвост, два крыла и две задние конечности. Кроме того, морфогенетическое поле обеспечивает лабильность, поскольку, если по каким-либо причинам доминирующая область морфогенетического поля разрушится, ее заменит соседняя. У кишечнополостных, например, головной отдел в норме образуется только из верхней части стебля. Но если головной отдел случайно разрушится, то из нижележащих тканей образуется новый.